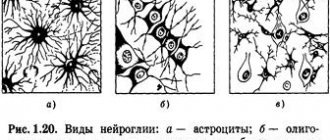
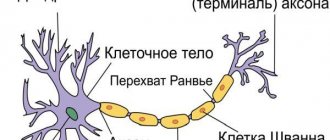
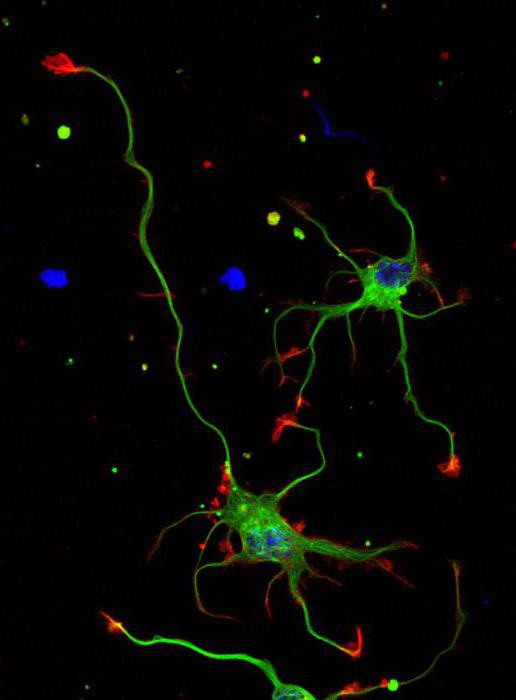
The nervous system consists of neurons (specific cells that have processes) and neuroglia (it fills the space between nerve cells in the central nervous system). The main difference between them is the direction of transmission of the nerve impulse. Dendrites are the receiving branches, along which the signal travels to the body of the neuron. Transmitting cells - axons - conduct a signal from the soma to the receiving cells. These can be not only neuron processes, but also muscles.
Differences between axons and dendrites
What is the difference between them? Let's consider.
- The neuron dendrite is shorter than the transmitting process.
- There is only one axon; there can be many receiving branches.
- The dendrites branch heavily, and the transmitting processes begin to separate towards the end, forming a synapse.
- Dendrites become thinner as they move away from the neuron body; the thickness of the axons remains virtually unchanged along their entire length.
- Axons are covered with a myelin sheath consisting of lipid and protein cells. It acts as an insulator and protects the process.
Because the nerve signal is transmitted as an electrical impulse, cells require insulation. Its functions are performed by the myelin sheath. It has tiny breaks that facilitate faster signal transmission. Dendrites are sheathless processes.
Definition
The medulla is a highly organized structure formed by nerve cells from which axons extend. Brain tissue is made up of nerve cells. Axon, translated from Greek, means “axis” - this is a process, an element of the medulla, which ensures interaction between cells of different types (neurons, cells of innervated organs), which is associated with subtle, precise control of the functioning of organs and systems. Functions of central nervous system tissue:
- Perceives irritations, converting them into impulses.
- Supports the transmission of impulses from the control parts of the brain to the executive organs.
- Forms a response to irritating influence.
- Provides interaction in the functioning of systems and organs, supports the integration of structural units of the body.
- Provides interaction between the body and the external environment.
According to the definition in biology, an axon is an elongated process along which impulses travel from the body of a neuron to other nerve cells and structural elements of all tissues of the body. During fetal development, brain tissue is formed from the neural plate. The edges of the plate bend, which leads to the formation of ridges and grooves. As a result of the closure of the edges of the ridges, a neural tube is formed - the basis of the central nervous system.
Differentiation of the cells forming the tube leads to the appearance of neuroblasts and spongioblasts. The former serve as the basis for the formation of neurons, the latter – for the formation of neuroglia. Neurons (anat.) are the main structural elements of the brain matter. They are characterized by the absence of a division function, which leads to a gradual decrease in their numbers. The body of a neuron consists of a nucleus and cytoplasm. Depending on the type of neurons, the geometric shape of the body changes, which can be round, oval, pyramidal and others.
The cytoskeleton, consisting of microtubules and neurofibrils, provides support and trophic function. The cytoskeleton maintains the shape of the neuron and ensures the transport of substances and organelles. Branches from the body include a single axon and multiple dendrites. The neuron axon has almost no branches and sometimes forms collateral (bypass) segments. The end segments (terminations) branch out and are called terminals.
The terminals are interconnected with the endings of other neurons and with cells that form the parenchyma (tissue) of working organs - muscles, glands. The number of dendrites varies from 1 to several. Thin branches of dendrites end in small spines, where the terminals of the axonal processes of many thousands of other cells are concentrated. Dendrites receive stimuli or action potentials from other cells and transmit them along fibers to the body of their neuron.
Axon growth depends on the structural features and vital activity of the neuron, which supports the function of feeding the process. For example, if an axonal trunk is cut, the segment connected to the body remains viable and continues to function, while the section that has lost connection with the body dies. Axons form nerves, which suggests a complex structural and morphological organization of the central nervous system.
Synapse
The place where contact occurs between branches of neurons or between an axon and a receiving cell (such as a muscle cell) is called a synapse. It may involve only one branch from each cell, but most often contact occurs between several processes. Each axon extension can contact a separate dendrite.
The signal at a synapse can be transmitted in two ways:
- Electric. This occurs only when the width of the synaptic cleft does not exceed 2 nm. Thanks to such a small gap, the impulse passes through it without being delayed.
- Chemical. Axons and dendrites come into contact due to the potential difference in the membrane of the transmitting process. On one side the particles have a positive charge, on the other - a negative one. This is due to different concentrations of potassium and sodium ions. The first are located inside the membrane, the second are outside.
When a charge passes through, the permeability of the membrane increases, and sodium enters the axon and potassium leaves it, restoring the potential.
Immediately after contact, the process becomes immune to signals, after 1 ms it is capable of transmitting strong impulses, and after 10 ms it returns to its original state.
Dendrites are the receiving side that transmits impulses from the axon to the body of the nerve cell.
Features characteristic of typical dendrites and axons
⇐ PreviousPage 3 of 14Next ⇒
| Dendrites | Axons |
| Several dendrites extend from the neuron body | A neuron has only one axon |
| Length rarely exceeds 700 µm | Length can reach 1 m |
| As you move away from the cell body, the diameter decreases rapidly | Diameter is maintained over a considerable distance |
| The branches formed as a result of division are localized near the body | Terminals are located far from the cell body |
| There are spines | No spines |
| Does not contain synaptic vesicles | Contains a large number of synaptic vesicles |
| Contains ribosomes | Ribosomes may be found in small numbers |
| Deprived of myelin sheath | Often surrounded by a myelin sheath |
The terminals of the dendrites of sensory neurons form sensory endings. The main function of dendrites is to receive information from other neurons. Dendrites conduct information to the cell body and then to the axon hillock.
Axon. Axons form nerve fibers that transmit information from neuron to neuron or to an effector organ. The collection of axons forms nerves.
It is generally accepted to divide axons into three categories: A, B and C. Fibers of groups A and B are myelinated, and C fibers are devoid of a myelin sheath. The diameter of group A fibers, which make up the majority of communications of the central nervous system, varies from 1 to 16 microns, and the speed of impulses is equal to their diameter multiplied by 6. Type A fibers are divided into Aa, Ab, Al, As. Ab, Al, As fibers have a smaller diameter than Aa fibers, a lower conduction speed and a longer action potential. Ab and As fibers are predominantly sensory fibers that conduct excitation from various receptors in the central nervous system. Al fibers are fibers that conduct excitation from spinal cord cells to intrafusal muscle fibers. B fibers are characteristic of preganglionic axons of the autonomic nervous system. The conduction speed is 3-18 m/s, the diameter is 1-3 µm, the duration of the action potential is 1-2 ms, there is no trace depolarization phase, but there is a long hyperpolarization phase (more than 100 ms). The diameter of C-fibers is from 0.3 to 1.3 microns, and the speed of impulses in them is slightly less than the diameter multiplied by 2 and equals 0.5-3 m/s. The duration of the action potential of these fibers is 2 ms, the negative trace potential is 50-80 ms, and the positive trace potential is 300-1000 ms. Most C fibers are postganglionic fibers of the autonomic nervous system. In myelinated axons, the speed of impulse conduction is higher than in unmyelinated axons.
The axon contains axoplasm. In large nerve cells, it accounts for about 99% of the entire cytoplasm of the neuron. The cytoplasm of axons contains microtubules, neurofilaments, mitochondria, agranular endoplasmic reticulum, vesicles and multivesicular bodies. In different parts of the axon, the quantitative relationships between these elements change significantly.
Axons, both myelinated and unmyelinated, have a sheath called the axolemma.
In the zone of synaptic contact, the membrane receives a number of additional cytoplasmic connections: dense projections, ribbons, subsynaptic network, etc.
The initial section of the axon (from its beginning to the place where it narrows to the diameter of the axon) is called the axon hillock. From this location and the appearance of the myelin sheath extends the initial segment of the axon. In unmyelinated fibers, this part of the fiber is difficult to determine, and some authors believe that the initial segment is characteristic only of those axons that are covered with a myelin sheath. It is absent, for example, in Purkinje cells in the cerebellum.
At the point of transition of the axon hillock into the initial segment of the axon, a characteristic electron-dense layer appears under the axolemma, consisting of granules and fibrils, 15 nm thick. This layer is not connected to the plasma membrane, but is separated from it by intervals of up to 8 nm.
In the initial segment, compared to the cell body, the number of ribosomes sharply decreases. The remaining components of the cytoplasm of the initial segment - neurofilaments, mitochondria, vesicles - move from the axon hillock here, without changing either in appearance or in relative position. Axo-axonal synapses are described at the initial segment of the axon.
The part of the axon covered with the myelin sheath has only its inherent functional properties, which are associated with the conduction of nerve impulses at high speed and without decrement (attenuation) over considerable distances. Myelin is a waste product of neuroglia. The proximal boundary of the myelinated axon is the beginning of the myelin sheath, and the distal boundary is the loss of it. This is followed by more or less long terminal sections of the axon. This part of the axon lacks granular endoplasmic reticulum and very rarely contains ribosomes. Both in the central parts of the nervous system and in the periphery, axons are surrounded by processes of glial cells.
The myelinated membrane has a complex structure. Its thickness varies from fractions to 10 microns or more. Each of the concentrically located plates consists of two outer dense layers forming the main dense line, and two light bimolecular layers of lipids separated by an intermediate osmiophilic line. The intermediate line of axons of the peripheral nervous system is the junction of the outer surfaces of the plasma membranes of the Schwann cell. Each axon is accompanied by a large number of Schwann cells. The place where Schwann cells border each other is devoid of myelin and is called the node of Ranvier. There is a direct relationship between the length of the interinterceptor section and the speed of nerve impulses.
The nodes of Ranvier constitute a complex structure of myelinated fibers and play an important functional role in conducting nerve excitation.
The length of the node of Ranvier of myelinated axons of peripheral nerves is in the range of 0.4-0.8 μm; in the central nervous system, the node of Ranvier reaches 14 μm. The length of the interceptions changes quite easily under the influence of various substances. In the area of the interceptions, in addition to the absence of the myelin sheath, significant changes in the structure of the nerve fiber are observed. The diameter of large axons, for example, is reduced by half, while small axons change less. The axolemma usually has irregular contours, and under it lies a layer of electron-dense substance. At the node of Ranvier there can be synaptic contacts both with dendrites adjacent to the axon (axo-dendritic) and with other axons.
Axon collaterals. With the help of collaterals, nerve impulses spread to a larger or smaller number of subsequent neurons.
Axons can divide dichotomously, as, for example, in granule cells of the cerebellum. The main type of axonal branching is very common (pyramidal cells of the cerebral cortex, basket cells of the cerebellum). Collaterals of pyramidal neurons can be recurrent, oblique and horizontal. The horizontal branches of the pyramids sometimes extend 1-2 mm, uniting the pyramidal and stellate neurons of their layer. From the horizontally spreading (in the transverse direction to the long axis of the gyrus of the brain) axon of the basket cell, numerous collaterals are formed, which end in plexuses on the bodies of large pyramidal cells. Such devices, as well as the endings on Renshaw cells in the spinal cord, are the substrate for the implementation of inhibition processes.
Axon collaterals can serve as a source of formation of closed neural circuits. Thus, in the cerebral cortex, all pyramidal neurons have collaterals that take part in intracortical connections. Due to the existence of collaterals, the safety of the neuron is ensured during the process of retrograde degeneration if the main branch of its axon is damaged.
Axon terminals. Terminals include the distal portions of axons. They lack a myelin sheath. The length of the terminals varies significantly. At the light-optical level, it is shown that the terminals can be either single and have the shape of a club, reticulate plate, ring, or multiple and resemble a brush, cup-shaped, mossy structure. The size of all these formations varies from 0.5 to 5 microns or more.
Thin axonal branches at points of contact with other nerve elements often have spindle-shaped or bead-shaped extensions. As electron microscopic studies have shown, it is in these areas that there are synaptic connections. The same terminal allows one axon to make contact with many neurons (for example, parallel fibers in the cerebral cortex) (Fig. 1.2).
Functions of nerve fibers
Spread of excitation in nerve fibers. Changes in membrane potential caused by electric current are divided into passive and active.
Passive, or electrotonic, changes in membrane potential are determined by the physical (electrical) parameters of both the membrane itself and the entire cell (fiber) as a whole.
Passive shifts in membrane potential occur when excitable formations are exposed to an electric current of any strength, shape or direction. However, if with hyperpolarizing (anodic) and weak depolarizing (cathodic) currents passive changes in potential can be observed in a pure (uncomplicated) form, then with close to the threshold and supra-threshold depolarizing stimuli they are accompanied by active potential shifts: local response and action potential associated with the changes ionic permeability of the membrane.
The passive properties of the membrane and the entire fiber as a whole largely determine the conditions for the occurrence and propagation of excitation in the nerve fiber.
Studies show that in a uniformly polarized, homogeneous section of a nerve fiber, changes in membrane potential when applying a rectangular impulse of a hyperpolarizing or weak depolarizing current increase exponentially:
,
where RC = τ
– membrane time constant, i.e.
the time during which the potential increases to 63% of its final value. When the current is turned off, the potential returns to its original level exponentially with the same time constant τ
. Such changes in membrane potential are usually called passive or electrotonic, in contrast to active ones associated with an increase or decrease in ionic conductivity of the membrane.
Similar changes are observed on spherical cells (on the soma). The description of a cylindrical cell, in particular an axon, is more complex. In this case, the inner conductor can no longer be considered equipotential along its entire length. The outer conductor can be considered equipotential due to an increase in the volume of extracellular fluid. The potential on such a membrane depends not only on the time the current is turned on, but also on the distance x
relative to the place where the current is applied:
,
where a
– radius of the fiber,
R
– resistivity of the axoplasm,
CM
and
RM
– capacitance and resistance per unit area of the membrane. The left side of the equation describes the current density through each point of the membrane, which is equal to the sum of the capacitive () and ohmic () current densities on the right side of the equation.
After a long time (much longer time constant t = RM CM
) after the pulse is turned on, the membrane capacitance will be fully charged and the capacitive current will become zero. The equation will take the form:
.
His solution:
,
where V0
– potential at the beginning of the cable (
x
= 0),
l
– fiber length constant.
The length constant characterizes the steepness of the potential attenuation along the fiber. The more l
, the further the signal travels along the fiber.
The speed of electrotonic propagation is proportional to twice the fiber length constant l
and inversely proportional to the time constant
t = RM CM
.
The value l
can be expressed through the membrane resistance
RM
, the resistance of the internal environment - axoplasm
Ri
and the fiber diameter
d
:
.
The cable properties of nerve fibers have a significant impact not only on the development of electrotonic potentials, but also on the nature of active responses - the threshold value, amplitude, slope of rise and duration of the action potential.
At present, it can be considered strictly proven that the conduction of the action potential (AP) along the nerve fiber is carried out with the help of local currents that arise between the excited and resting sections of the membrane. The local current changes the value of the resting membrane potential in the resting region to a critical level of depolarization, which is the cause of the action potential.
Numerous studies have shown that the conduction speed is proportional to the constant fiber length l
and is inversely proportional to the membrane time constant
t
(Chailokhyan L.M., 1962).
Since in non-pulp nerve fibers l
is proportional to the square root of the fiber diameter
,
the conduction speed, other things being equal, is also proportional to the square root of the fiber diameter.
In myelinated nerve fibers, conduction occurs saltatory - from the node of Ranvier to the node of Ranvier. The length of the inter-intercept section is approximately proportional to the fiber diameter, so the conduction velocity in these fibers is not proportional to the square root of the fiber diameter, but simply to its diameter.
It is generally accepted that the speed of implementation depends on the value of the so-called safety factor (warranty) F
, i.e. the ratio of the amplitude of the propagating AP to the threshold potential. The threshold potential is the amount by which the membrane potential must be changed to achieve a critical level of depolarization.
,
where Vs
– AP amplitude,
Vt
– threshold potential.
When Ф = Vt
there is no spread of excitation. For a crab axon this ratio is 7.
It was shown that the threshold potential Vt
is closely dependent on the sensitivity of the sodium permeability system of the membrane to depolarization.
The higher this sensitivity, i.e. the greater the increase in PNa
and, accordingly, the incoming sodium current
INa
at a given potential shift, the lower the threshold, and vice versa. Changing the state of the potassium permeability system has virtually no effect on the threshold potential. In the same way, the conductivity of “leakage” currents has very little effect on the threshold potential. At a constant resting potential, the safety factor should increase with effects on the nerve fiber that increase the sensitivity of the sodium system to depolarization, for example, reducing the concentration of calcium ions in the environment. A significant decrease in the safety factor is caused by agents that enhance the initial inactivation of the sodium system or reduce sodium conductance, since in this case the amplitude of the action potential decreases and the threshold potential increases. Such changes in the conduction of excitation were observed by Tasaki (1957) and other researchers when the nerve fiber was exposed to anesthetics and drugs in small concentrations, insufficient to completely suppress the action potential.
The level of resting potential has a complex influence on the safety factor. Short-term subthreshold depolarization of the membrane, which does not significantly change the critical potential and amplitude of the action potential, increases the safety factor, since Vt = Eo – Ek
. With strong depolarization, the spike amplitude decreases, the critical potential increases, and therefore the safety factor decreases.
Along with the safety factor, the steepness of the ascending phase of the propagating action potential has a significant influence on the speed of excitation. The slope of this phase depends on both the passive and active properties of the membrane.
Approximately 1/3 of the ascending phase of the propagating AP is associated with passive depolarization of the nerve fiber membrane by local circuit current. The rate of this depolarization at a given local current strength is determined by the membrane time constant t = RM CM
. The smaller this value, the faster the depolarization increases and, therefore, the steeper the spike rises. Inactivation of the sodium system, or decrease in sodium permeability (active properties of the membrane), sharply reduces the slope of the ascending phase. Thus, for most impacts, changes in the rate of increase in the ascending phase of the PD coincide in their direction with changes in the safety factor.
According to the theory of local currents, the amplitude of the propagating action potential Vs
, in contrast to the membrane spike, depends not only on the EMF of the excited membrane
E
, but also on the ratio of the input resistances of the excited
R1
and unexcited (load resistance
R2
) sections of the fiber:
. (1)
The higher the ratio, the more the amplitude of the propagating PD approaches the value E
, the higher the safety factor, and vice versa. From which it follows that a decrease in membrane resistance (increase in its ionic conductivity) during critical depolarization not only leads to the occurrence of a spike, but also contributes to an increase in the safety factor, and therefore the conduction speed.
From formula (1) it is clear that when excitation is carried out along geometrically inhomogeneous excitable conductors, the amplitude of the propagating spike should significantly depend on how close the currently excited section of the fiber is to the place of its branching or expansion.
When a nerve fiber expands, for example, at the point of its transition into the cell body or in the area of axon branching, the total cross-sectional area of the fibers and the total area of their membrane increases, and therefore R2
falls.
A decrease in R2
reduces the safety factor and, accordingly, the speed of conduction.
Under some conditions, a decrease in R2
can lead to complete blocking of a nerve impulse.
Calculations have shown that the action potential easily passes threefold expansion, with difficulty fivefold, and is completely blocked at sixfold. The reason for the development of the block is a sharp decrease in the amplitude of the propagating PD near the region of fiber expansion.
Trophic function of nerve fibers. Afferent and efferent fibers have a trophic function.
Afferent nerves have two neurotrophic, non-impulse functions. It is possible to distinguish between a direct morphogenetic and trophic effect on peripheral organs and a regulatory function with feedback, probably dependent on intra-axonal centripetal impulses. Neurotrophic morphogenetic influence is proven by the presence of: a) dependence of the structure of taste buds on taste nerves; b) stimulating the regeneration of limbs in amphibians by sensory nerves through a specific, growth-stimulating substance of a non-mediator nature; c) differentiation and maintenance of receptors. After deafferentation, trophic disorders develop in some organs. The primary “trophic” neuron for a muscle is a motor neuron. We must also not forget that efferent adrenergic fibers pass through all nerves, in which neurosecrets (catecholamines) are transported by axoplasmic current to peripheral organs.
Axonal transport. Two systems of axonal transport have been described: slow, with a speed of 1-3 mm/day, and fast, with a speed of approximately 400 mm/day.
Axonal transport maintains the continuity of the axon and synaptic membranes and restores proteins, glycoproteins, enzymes and other substances that disappear during local breakdown, exocytosis into the synaptic cleft and retrograde migration to the neuron. All this occurs due to fast transport, which is not influenced by excitation processes. Transport continues after blockade of action potentials and does not increase with increased nerve activity. Axonal transport occurs in both directions; the centripetal current apparently controls the synthesis of proteins in the neuron and also plays the role of a “signal” for chromatolysis after axotomy. Various substances, enzymes, transmitters and macromolecules move in the axon at different speeds.
Axoplasmic transport can be recorded by the accumulation of substances after disruption of axonal continuity and by monitoring the progress of labeled compounds after they are introduced into the neuron.
Proteins synthesized in the cell body, synaptic mediator substances and low molecular weight factors descend along the axon to the nerve terminal along with cellular organelles, in particular mitochondria. For most substances and organelles, retrograde transport has been detected (along the axon to the cell body): polio virus, herpes virus, tetanus toxin, as well as enzymes - horseradish peroxidase, which is widely used in neuroanatomy as a marker. Retrograde transport appears to be the main factor in the regulation of protein synthesis in the cell. After transection of the axon, chromatolysis begins in the soma a few days later, which indicates a violation of protein synthesis. Rapid axonal transport depends on an adequate supply of metabolic energy. The possibility of transport is created by microtubules with a diameter of 25 microns, consisting of the protein tubulin, and some neurofibrils, consisting of the protein actin, forming transport threads. Transport threads slide along microtubules. At the same time, they interact with the protrusions of microtubules, ATP is broken down, which provides energy for transport. Large proteins are transported more slowly. But it is believed that the transport mechanism itself is not slower, but substances from time to time enter cellular compartments that are not involved in transport. Slow current also appears to be related to axonal growth. Axoplasmic flow is stopped by colchicine, which is explained by the effect of this substance on microtubules.
Physiology of synapses
Synapse (from the Greek synapsis) means a connection; a connection is a specialized zone of contact between neurons or neurons and other excitable formations, ensuring the transfer of excitation with the preservation, change or disappearance of its information value. This term was proposed by C. Sherrington (1897) to denote the functional contact between neurons. To be fair, it should be noted that back in the 60s of the 19th century I.M. Sechenov emphasized that the origin of even the simplest reflexes cannot be explained outside of intercellular communication.
Synapses are distinguished: 1) by their location; 2) by the method of signal transmission.
1) By location, synapses are distinguished as central and peripheral. Central synapses are synapses that make contact between neurons in the central nervous system. These include axo-axonal synapses, axo-dendritic, axo-somatic, dendro-dendritic (discovered histologically; the functional significance is not entirely clear). Central synapses are also classified according to the sign of their action - excitatory and inhibitory. In addition, it is common to divide synapses according to the mediator (transmitter) that mediates: adrenergic synapses, cholinergic synapses, etc.
Peripheral synapses include neuromuscular synapses, synapses of the autonomic ganglia (synapses formed by preganglionic and postganglionic fibers).
2) According to the method of transmission, synapses are classified as chemical and electrical.
All these formations are characterized by the presence of a presynaptic membrane, a synaptic cleft (10-50 nm), and a postsynaptic membrane. The presynaptic membrane is the membrane of the presynaptic end of a neuron process (most often an axon).
In humans and higher vertebrates, chemical synapses are most widespread. Chemical synapses at the presynaptic terminal contain vesicles with a mediator, a chemical transmitter. The width of the synaptic cleft is on average 20 nm. The postsynaptic membrane contains receptors for this mediator and enzymes that destroy this mediator. Thus, the postsynaptic membrane is the receptor part of the synapse; it can be a specifically differentiated area of the dendrite, the body of the neuron and its axon.
The electrical synapse does not produce a transmitter. The synaptic cleft is somewhat smaller than that of a chemical synapse (2-4 nm). In the synaptic cleft between the pre- and postsynaptic membranes there are protein bridges-channels 1-2 nm wide, where ions and small molecules move. This contributes to a lower resistance of the postsynaptic membrane than that of the presynaptic membrane. Therefore, excitation from the presynaptic membrane to the postsynaptic membrane in electrical synapses is transmitted electrically, i.e. ephaptic transmission occurs. Unlike chemical synapses, electrical synapses are distinguished by a higher speed of excitation, high reliability of transmission, and the possibility of bilateral conduction.
Electrical synapses were found in rats in the vestibular nucleus of the medulla oblongata, in the structures of the respiratory center of the medulla oblongata (their role in the mechanisms of automatic rhythmogenesis of breathing is discussed); in cats, electrical synapses are found between neurons of the inferior olives, in the structures of the thalamus, between photoreceptors of the retina and horizontal cells in fish, etc.
But still, chemical synapses became most widespread in the process of evolution. This is due to a number of properties of these formations, which are of great importance in organizing the activity of the nervous system (Fig. 1.4).
Rice. 1.4.
Synapse (picture taken from the book: Brain / edited by P.V. Simonov. M.: Mir, 1984)
⇐ Previous3Next ⇒
Recommended pages:
Functioning of the nervous system
The normal functioning of the nervous system depends on impulse transmission and chemical processes at the synapse. The creation of neural connections plays an equally important role. The ability to learn is present in humans precisely because of the body's ability to form new connections between neurons.
Any new action at the learning stage requires constant control by the brain. As it is mastered, new neural connections are formed, and over time the action begins to be performed automatically (for example, the ability to walk).
Dendrites are transmission fibers that make up approximately one third of all nervous tissue in the body. Through their interaction with axons, humans have the ability to learn.